












本期解读

一、研究背景
人参含有多种黄酮醇苷,具有多种生物活性,然而关于人参中黄酮糖基化的报道很少。在人参中,人参皂苷和山柰酚3- O-葡萄糖苷通常随着种植年限在叶片中积累。探索人参中黄酮醇糖基化的过程,有助于揭示人参在生物过程中的调节作用。

二、研究结果
1.人参中黄酮醇糖苷类物质的时空分布特征
采用UPLC-MS-QqQ对不同栽培年份(2、3、4年)人参叶、茎、根中的黄酮类化合物进行了分析(图1A),共鉴定出六种黄酮醇化合物:山奈酚(K)、山奈酚3-葡萄糖苷(K3Glc)、山奈酚3-O-葡萄糖基-(1-2)-半乳糖苷(K3GalGlc、人参皂苷)、槲皮素(Q)、槲皮素3-葡萄糖苷(Q3Glc、异槲皮素)和槲皮素3-O-葡萄糖基-(1-6)-鼠李糖苷(芦丁)。黄酮类化合物主要积累在叶片和茎中,其次是根(图1B, C)。随着年份的增加,叶和根中总黄酮含量增加,而茎中总黄酮含量下降,芦丁和山奈酚3-葡萄糖苷随着培养年限的增加而显著下降。根中山奈酚3-O-葡萄糖苷含量增加较小,但在4年叶中增加显著,此外,类黄酮单葡萄糖苷山奈酚3-O-葡萄糖苷在2岁茎中的含量高于4岁时的叶片含量。
4岁龄的叶片中山奈酚3-O-葡萄糖苷含量为148μg/g DW,山奈酚二糖苷(泛烯苷)含量高达1584μg/g DW。有趣的是,人参根和3年或4年的茎中山奈酚和槲皮素葡萄糖苷的含量均很低,但在2年的茎中发现山奈酚3-O-葡萄糖苷(264.9μg/g DW)和芦丁(442.9μg/g DW)的含量较高,可能是因为幼茎也能合成黄酮类化合物。山奈酚和被测组织中槲皮素含量均较低,但山奈酚和槲皮素的糖基化产物以积累为主,说明糖基化在人参类黄酮稳定性中起着重要作用。
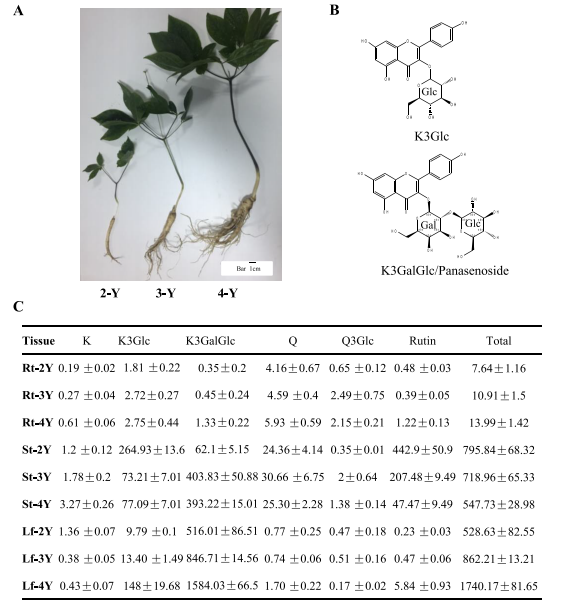
图1 人参中黄酮醇苷的时空分布图。(A)不同栽培年份的人参性状;(B)泛烯苷(K3GalGlc)和山奈酚3-O-葡萄糖苷(K3Glc)的结构;(C)不同栽培年份人参不同组织中类黄酮含量(μg/g DW)。
2.山奈酚3-O-葡萄糖苷的差异蛋白质组学及PgUGTs的鉴定
对2岁龄的人参植株中叶和根进行定量DIA分析,共鉴定到4535个蛋白质,结合225个注释的UGT蛋白,筛选了19个PgUGTs,以评价其在黄酮醇苷形成中的作用。以UDP-葡萄糖为糖供体,重组PgUGT7和PgUGT12向山奈酚的3-OH中添加葡萄糖。以山奈酚为底物的PgUGT7和PgUGT12酶解产物的保留时间和MS光谱与标准山奈酚3-O-葡萄糖苷相同(图2B, C)。蛋白质组学数据(图2A)显示,叶片中PgUGT7和PgUGT12的丰度分别是根的1.7倍和5.4倍,与代谢组学数据(图1C)一致。
山奈酚苷是一种山奈酚二苷(山奈酚3-O-糖基-(1-2)半乳糖苷),可由山奈酚3-O-半乳糖基转移酶和山奈酚3-O-半乳糖苷糖基转移酶产生,分别促进山奈酚3-O-半乳糖苷和泛烯苷的形成。以UDP-半乳糖/葡萄糖作为糖供体,以山奈酚或山奈酚3-O-半乳糖苷作为底物,对所有候选PgUGTs进行了测试。系统发育分析显示PgUGT7和PgUGT12在黄酮醇的多个位点与糖基化的进化群聚集在一起,例如UGT79家族(图2D),PgUGT12对山奈酚和山奈酚3-O-半乳糖苷的酶促能力与其在进化树中的位置一致,接近UGT79家族。
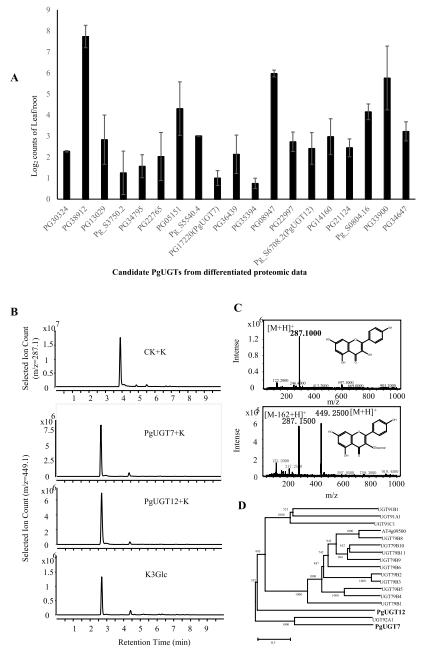
图2 PgUGTs差异蛋白质组学。 (A) 19个PgUGTs蛋白在叶片向根的表达水平;(B) 从PgUGT7和PgUGT12酶产物的UPLC-MS中提取的离子;(C)山奈酚和山奈酚3-O-葡萄糖苷(K3Glc)的质谱图;(D) PgUGT7、PgUGT12和AtUGTs的分子系统发育树。
3.系统发育分析筛选候选PgUGTs
利用系统发育分析进一步筛选人参基因组中注释的225个PgUGTs。从UGT71、UGT73、UGT74、UGT74、UGT75、UGT76、UGT79、UGT84、UGT85、UGT91和UGT94家族中选择了34个候选PgUGTs进行进一步分析,包括已报道的与其他物种中类黄酮相关的UGTs(图3A)。已报道的UGTs系统发育分析表明,49个UGTs可分成5个主要分支,为候选UGTs可能的酶功能提供证据(图3B)。其中,以ZmF3GT为代表的第一个分支由8个UGT组成,它们催化黄酮类化合物3-OH位点的糖基化;第二个分支包含5个糖基化类黄酮5-OH位点的UGTs,说明簇中的PgUGTs可能催化黄酮类化合物5-OH的糖基化;PgUGT73A18与8个UGTs聚集在第三个分支中,这些UGTs对黄酮类化合物的7-OH位点进行糖基化;PgUGT71A27、PgUGT71A40和PgUGT71A41形成一个新分支,其中PgUGT71A27可以催化PPT型人参皂苷的糖基化,因此,可以猜测PgUGT71A40和PgUGT71A41参与了人参皂苷的糖基化;分支形成簇由13个UGTs组成,它们能催化多步糖基化合成黄酮类化合物,该集群包括来自UGT79、UGT91和UGT94家族的七个候选PgUGT,包括上述确定的候选UGT94Q4。因此,选择分支形成簇的PgUGT91S2/V1、PgUGT79B38和PgUGT94X1/W1/Q2作为下一步酶实验的重点。
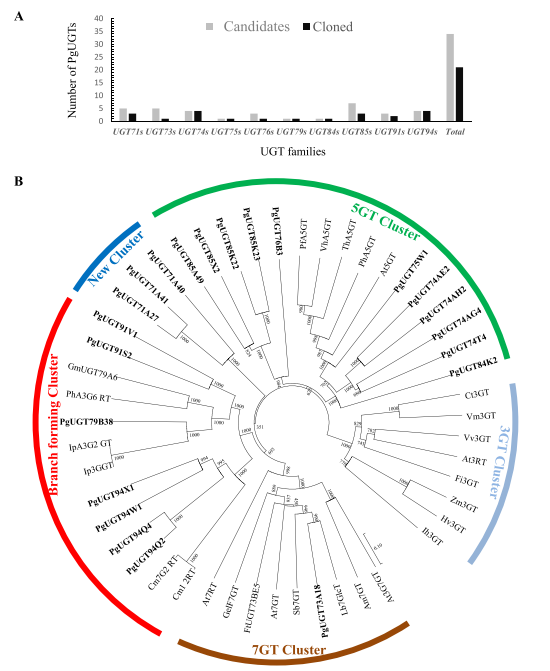
图3 基于氨基酸序列的参与类黄酮糖基化的PgUGT的分子系统发育树。(A)从不同UGT家族筛选的34个候选PgUGTs;(B)用ClustalX2比对多个序列,并使用MEGA7.0构建树。
4.候选PgUGTs的酶活性
利用原核系统鉴定了22个重组PgUGT蛋白的功能来研究黄酮类化合物的二次糖基化。以UDP-葡萄糖或UDP-半乳糖为糖基供体,以山奈酚、槲皮素、山奈酚3-O-葡萄糖苷和槲皮素3-O-葡萄糖苷为底物进行酶活性测定。结果证明,UGT75W1、UGT74T1和UGT73A18可催化槲皮素的羟基葡萄糖基化和山奈酚3-O-葡萄糖的半乳糖基化,UGT75W1和UGT73A18可催化山奈酚的葡萄糖基化(图4A-C)。通过对酶产物的分析表明,重组UGT75W1、UGT74T4和UGT73A18产生了含有山奈酚和槲皮素的新产物(图4D)。系统发育分析表明,UGT73A18属于5-O-GT分支,但酶活性分析表明,它能产生多种类黄酮苷,包括5-O-糖苷和3-O-糖苷。UGT75W1和UGT74T4属于7-O-GT分支,在酶活性测定中,两者都能使7-OH位点糖基化;此外,UGT74T4还可以催化微量5-O-糖苷的形成。以上结果表明这三种糖基转移酶具有底物特异性和区域选择性。
虽然UGT73A18、UGT74T4和UGT75W1均可催化黄酮醇糖基化,但每一种都有自己独特的催化特性。UGT73A18和UGT75W1催化槲皮素和山奈酚转化为相应的3-O-葡萄糖苷衍生物,而UGT74T4催化黄酮醇形成相关的7-O-葡萄糖苷和5-葡萄糖苷。酶动力学实验表明,当使用UDP半乳糖作为糖供体时,UGT75W1、UGT73A18和UGT74T4对山奈酚3-O-葡萄糖的酶活性不同(图5A)。三种PgUGTs在底物特异性(对黄酮醇的偏好)和多样性(对不同黄酮醇的亲和力)方面的相似性可能主要是由于它们的结构差异。
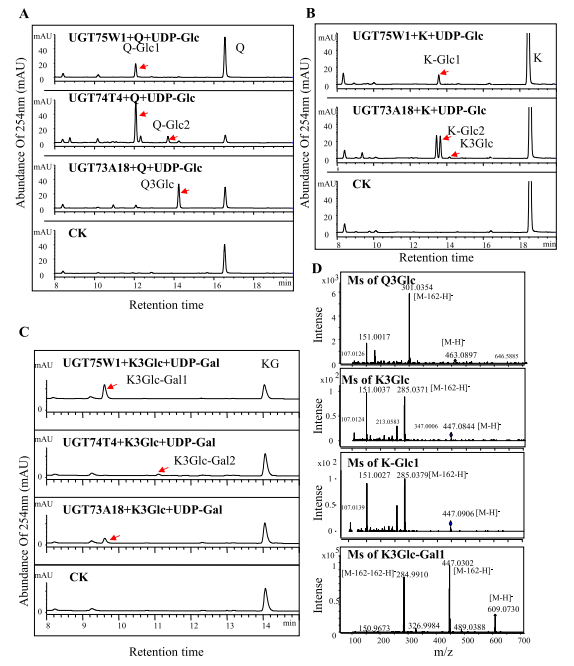
图4 黄酮醇重组PgUGT酶促产物的鉴定。含有槲皮素(A)、山奈酚(B)、山奈酚3-O-葡萄糖苷(K3Glc)和UDP-半乳糖(UDP-Gal)(C)和重组UGT75W1、UGT74T4以及UGT73A18的酶产物的代表性HPLC色谱图;(D)重组PgUGTs酶产物的质谱。
5.PgUGTs的分子建模与对接
通过分子对接模型得知,UGT73A18中包括Gly15、Ser286、Trp344、Gln347和Ser367在内的残基可以与山奈酚形成氢键。UGT75W1和山奈酚3-O-葡萄糖苷之间形成氢键的关键位点包括Gln15、Thr254和Leu255,而UGT75W1和UDP半乳糖之间形成氢键的关键位点包括Ser278、Trp335、Gln338、Val339(图5A-E)。UGT73A18和UGT74T4与UDP半乳糖和山奈酚3-O-葡萄糖苷的对接结果与UGT75W1不同,与UDP半乳糖形成氢键的残基主要位于UGT73A18和UGT74T4的N-末端和环中,表明UGT75W1中接近中性粒的残基可以与UDP葡萄糖结合,氨基酸位点Ser278、Trp335、Gln338和Val339可能是UGT75W1和山奈酚3-葡萄糖苷通过分子对接有效结合的关键位点。
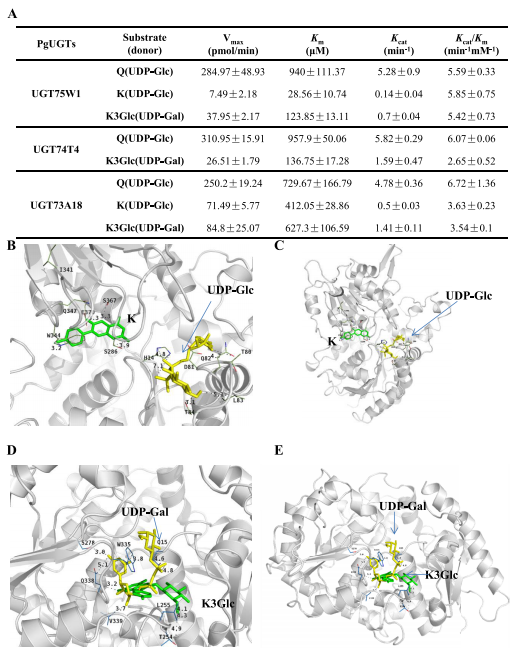
图5 PgUGTs的特性、同系物模型及与黄酮类化合物的对接。(A) 重组PgUGTs的代表性动力学参数;(B, C) UGT73A18与UDP-葡萄糖(UDP-Glc)和山奈酚(K)对接的的代表性结合域和整体视图;(D, E) UGT75W1与UDP-半乳糖(UDP-Gal)和山奈酚3-O-葡萄糖(K3Glc)对接的代表性结合域和整体视图。
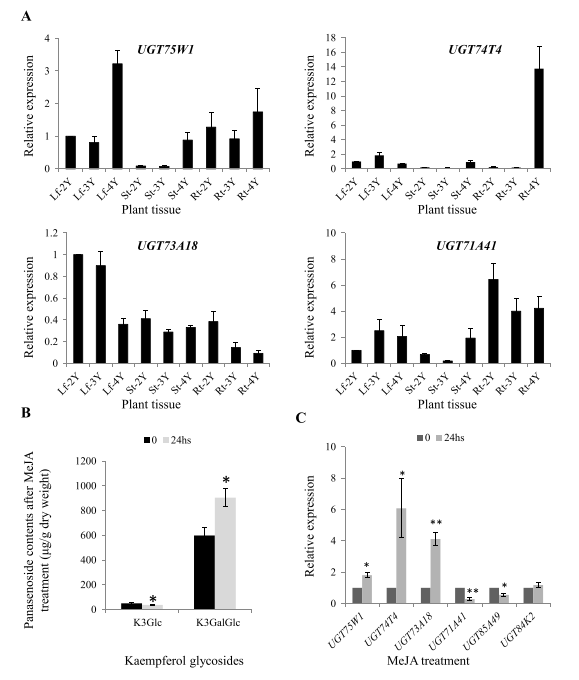
6. MeJA处理后植物中PgUGTs的表达谱
利用UGT73A18、UGT74T4和UGT75W1的转录表达谱结合它们的酶活性和组织表达谱(图6A),结果表明UGT73A18可能参与了叶和茎中黄酮醇苷的形成,UGT73A18、UGT74T4和UGT75W1可使3-O-葡萄糖苷半乳糖基化,因此根据UGT74T4和UGT75W1的表达模式,人参根或叶中可能存在至少两种山奈酚二糖苷。此外,MeJA处理结果(图6B-C)显示山奈酚3-O-葡萄糖苷的含量下降,表明这些PgUGTs在体内以山奈酚3-O-葡萄糖苷为底物催化山奈酚二糖苷的形成。MeJA处理诱导了UGT73A18、UGT74T4和UGT75W1的表达,表明其可能有助于不利条件下黄酮醇葡萄糖苷的产生,同时MeJA处理可诱导幼苗产生黄酮醇葡萄糖苷。
三、结论
研究证明T2DM-DN患者补体系统过度激活,表现为补体蛋白尿排泄异常。尿蛋白CFAH的较高丰度和DAF的较低丰度与进展为ESRD较高的风险密切相关。这些结果表明,补体途径的过度激活可能与进展为ESRD的更高风险有关。因此,治疗性靶向补体途径可以缓解DN的进展。
文章链接:
https://doi.org/10.1007/s40618-021-01596-3.
参考文献:
Zhao L., Zhang Y., Liu F. et al. Urinary complement proteins and risk of end-stage renal disease: quantitative urinary proteomics in patients with type 2 diabetes and biopsy-proven diabetic nephropathy[J]. J Endocrinol Invest, 2021, 44: 2709-2723.
编辑丨Aaliyah
审核丨Tao Li

