












病原体攻击可以增加植物活性氧(ROS)的水平,活性氧作为信号分子激活植物防御机制,阐明这些过程对于理解植物防御响应中的氧化还原信号通路至关重要。
华中农业大学在《Plant Communications》期刊发布题为"Comparative oxidation proteomics analyses suggest redox regulation of cytosolic translation in rice leaves upon Magnaporthe oryzae infection"的文章,报道了其通过氧化蛋白质组学揭示水稻通过核糖体蛋白半胱氨酸氧化响应稻瘟菌侵染。
研究背景
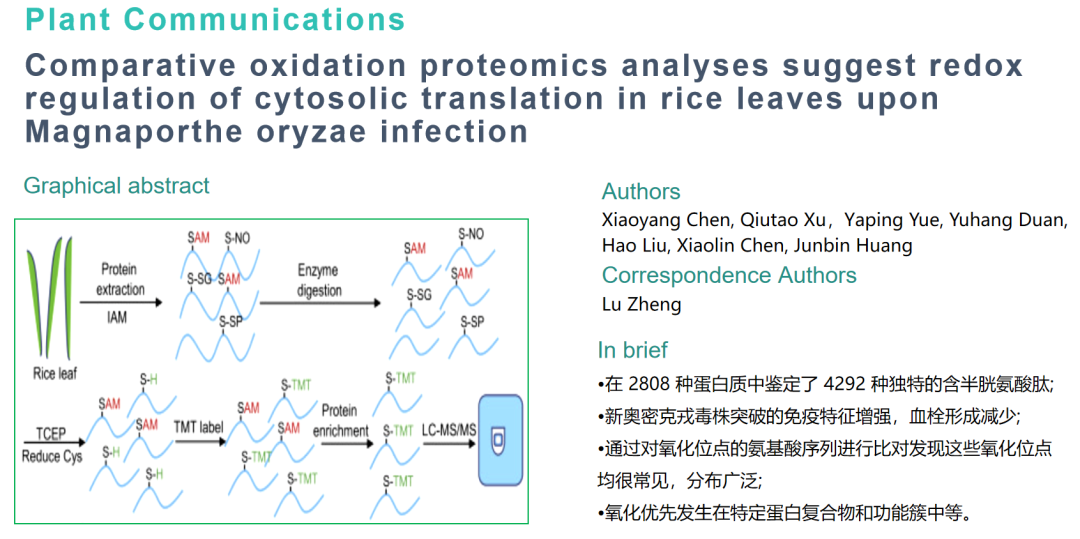
蛋白组翻译后修饰 (PTM) 广泛参与植物对病原菌侵染的调控,特别是精细调控,氧化还原调控是涉及转录、代谢过程、表观遗传过程和细胞信号传导的一般和基本机制,氧化还原调控通常发生在半胱氨酸残基的巯基部分,包括巯基/二硫过渡、s -谷胱甘肽酰化(S-SG)、亚砜化、s -亚硝基化(S-NO)、磺化和亚砜化。
作为世界一半以上人口的主食,水稻(水稻)对全球粮食安全至关重要,丝状真菌 Magnaporthe oryzae 引起稻瘟病,高度破坏栽培稻病害,威胁着世界各地的粮食生产 。M. oryzae 是一种模式生物,用于研究植物病原真菌的生理和致病分子机制,复杂的生理氧化还原平衡对于米分枝杆菌和水稻之间的相互作用至关重要 ,在这种相互作用的早期阶段,水稻会产生大量的ROS来帮助抵抗感染,从而导致蛋白氧化,进而改变它们的结构和分子功能,然而,ROS在植物细胞中信号传导和特定靶标调控中的确切作用在很大程度上是未知的。为了解决这些问题,作者使用串联质谱标签 (TMT) 蛋白质组学来表征稻米分枝杆菌感染期间水稻叶片细胞中蛋白质氧化水平的动态变化。
技术手段
实验材料:接种M. oryzae24h后的水稻叶片,喷洒H2O2的水稻叶片,正常的水稻叶片;
研究方法:TMT蛋白组、氧化修饰组、Ribo-seq。
研究内容
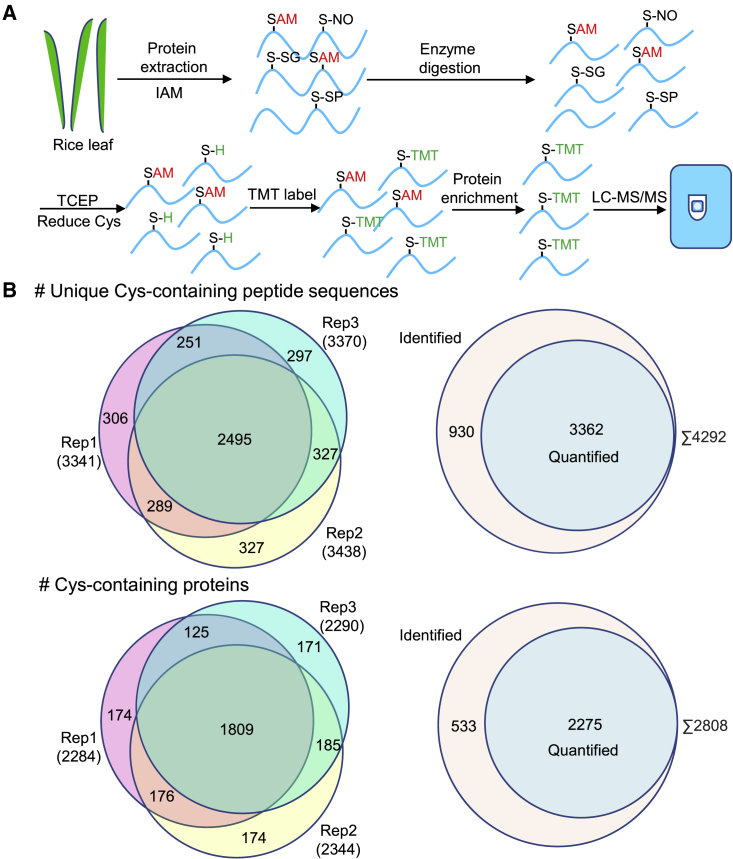
在2808 种蛋白质中共鉴定了 4292 种独特的含半胱氨酸肽
作者在 2808 种蛋白质中鉴定了 4292 种独特的含半胱氨酸肽,其中有2275 种蛋白质的 3362 个位点在三个生物学重复中的至少两个中被定量,这3362 个位点被认为是发现的位点总数,并做进一步分析,在 2275 种氧化蛋白中,1563 种只有一个氧化位点,696 种有 2-5 个氧化位点,16 种有 6 个或更多氧化位点,鉴定到氧化程度最高的蛋白质是生长素转运蛋白(由 Os09g0247700 编码),且通过分析发现,这些氧化位点很常见,分布广泛,且最常发生在极性较强的氨基酸附近。
氧化蛋白的亚细胞定位多位于叶绿体
为了确定水稻叶片中氧化蛋白的亚细胞定位,作者对这些蛋白进行了亚细胞定位预测,结果显示这2275种氧化蛋白,大多定位于叶绿体(41.8%),细胞质(22.6%)或细胞核(14.2%),作者还分析了不同修饰位点数量的氧化蛋白的亚细胞定位。在细胞核和线粒体中,具有5个以上修饰位点的蛋白质比例低于具有1个修饰位点的蛋白质比例,而在细胞质膜中观察到相反的趋势,氧化蛋白亚细胞定位的多样性表明氧化可能在多种细胞过程中起关键作用。氧化蛋白间的互作网络图(PPI)显示,氧化优先发生在特定的蛋白质复合物或功能簇中。GO通路分析显示,在生物过程类别中,氧化蛋白主要富集于细胞过程(41.3%)和代谢过程(32.8%),许多氧化蛋白与对刺激、信号传导和免疫系统的反应有关;在分子功能类别中,氧化蛋白的催化活性(57.9%)、结合活性(27.4%)、结构分子活性(4.8%)、分子功能调节活性(1.9%)和翻译调节活性(1.6%)富集;在细胞成分类别中,氧化蛋白主要富集在细胞解剖实体(78.4%)和含蛋白质复合物(21.6%)中。进一步的通路富集分析显示,基因表达、翻译、解毒、对应激的反应、代谢过程和生物合成过程显着富集;表明蛋白质氧化可能在正常条件下在这些代谢途径中具有重要的调节作用,这极大增加了氧化修饰的认识。
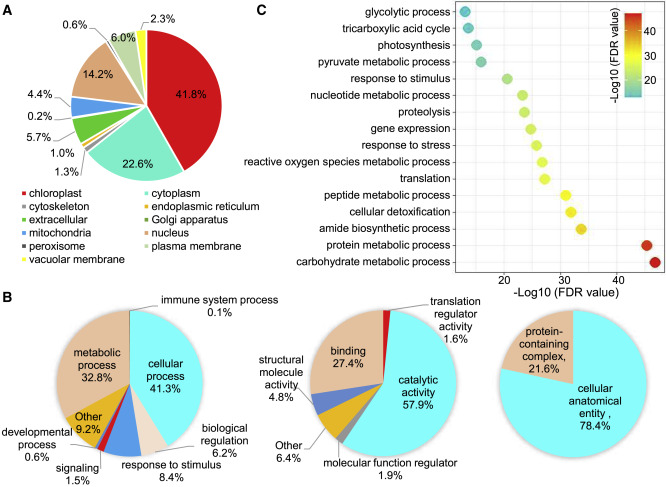
图1 水稻叶片的氧化位点分析
M. oryzae侵染的蛋白质氧化修饰组学分析
为了阐明氧化修饰的潜在功能,作者对M. oryzae侵染的叶片和对照组叶片进行氧化修饰组学分析。发现M. oryzae侵染组中,438种蛋白上的512个位点的氧化修饰水平明显上升,M. oryzae 处理导致的修饰水平改变与H2O2处理的叶片中修饰改变趋势一致,这表明M. oryzae侵染叶片中蛋白质的高氧化修饰水平是由ROS爆发引起。亚细胞定位分析表明侵染过程中超氧化蛋白的功能,侵染后细胞质中氧化修饰蛋白明显增加,因此细胞质中的氧化修饰蛋白可能是M. oryzae 侵染应答的关键。GO富集分析表明,氧化修饰蛋白在翻译、肽生物合成过程和肽代谢过程中富集,这表明氧化修饰可能参与M. oryzae 侵染期间水稻翻译过程的调节。
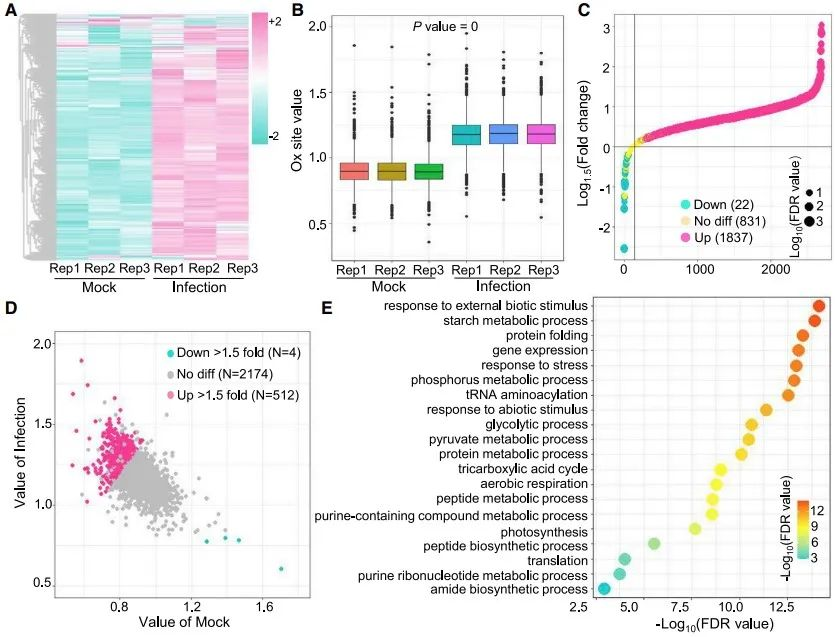
图2 M. oryzae 侵染后水稻叶片蛋白质氧化修饰水平增加
M. oryzae感染后核糖体蛋白氧化修饰水平增加,可促进胞质核糖体翻译
分析发现M. oryzae感染后叶片中33种核糖体蛋白和17种翻译因子的氧化修饰水平上升,说明M. oryzae感染后核糖体蛋白氧化修饰水平增加。先前的研究表明,核糖体蛋白的乙酰化、磷酸化和泛素化等 PTM 会影响核糖体功能 ,为了确定感染样本中核糖体蛋白氧化增加是否会改变核糖体翻译过程,作者进行了核糖体印记测序(Ribo-seq)来分析模拟样本和正常样本之间核糖体翻译的差异,分析结果显示,核糖体蛋白氧化修饰水平的增加促进了大量参与防卫反应的mRNA翻译效率,进而增强了水稻的免疫能力。也说明了植物细胞除转录调控外,还通过翻译调控来快速响应病原体入侵植物的变化。为了进一步验证该假设,作者还分析了侵染叶片和正常叶片的蛋白质组学数据,结果显示侵染叶片的总体蛋白质水平高于正常叶片,300种蛋白质水平显著上调,进一步证实了该假设。
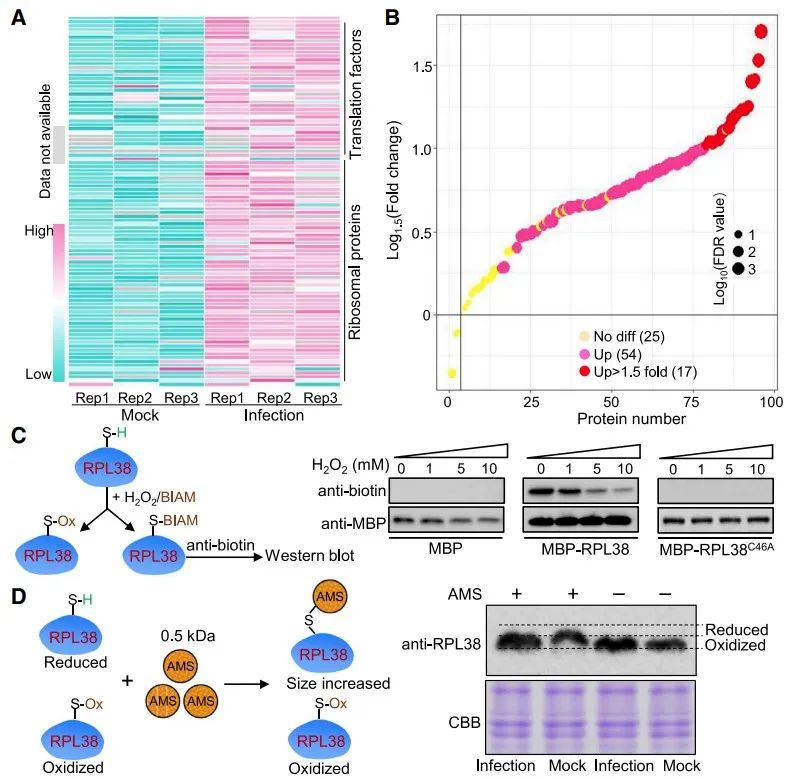
图3 M. oryzae感染后水稻叶片核糖体蛋白氧化修饰水平增加
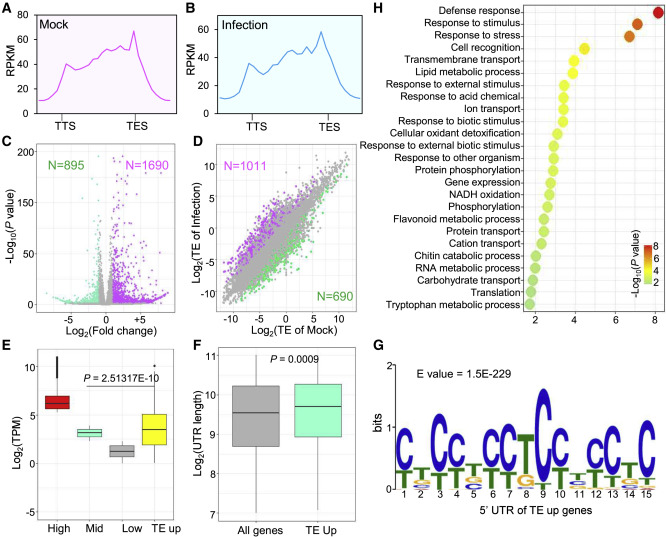
图4 M. oryzae感染后促进促进胞质核糖体翻译翻译水平增加
核糖体蛋白过度氧化促进胞质溶胶中蛋白质翻译
作者分析了来自感染和模拟叶子的蛋白质组学数据。两组之间的比较分析显示,感染组的总体蛋白质组学水平高于对照组。进一步比较感染叶与对照叶的数据分析显示,300种蛋白质显示出较高的蛋白质水平(>1.2倍,FDR<0.05),而只有100种蛋白质水平较低(>1.2倍,FDR<0.05)。此外,高翻译效率的1011 mRNA序列对应的蛋白水平在侵染叶片中显著高于对照叶片,支持了核糖体蛋白氧化增加导致翻译效率升高的假设,显示丰度显著增加/减少的蛋白质的免疫印迹进一步支持了这一假设。
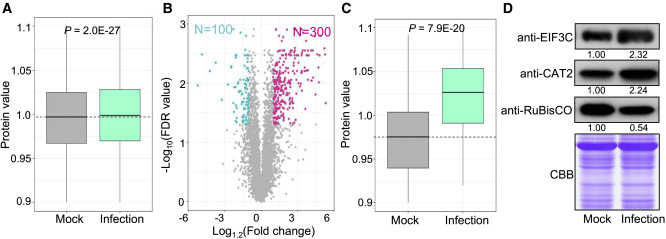
图5 水稻叶片M. oryzae感染组与正常对照组的蛋白组分析
研究结论
总而言之,该项研究通过TMT蛋白组学、氧化修饰组学、Ribo-seq分析发现病原体感染期间反应性氧化物水平升高导致包括代谢酶、蛋白激酶和转录因子等蛋白质氧化,还发现核糖体蛋白也被植物与病原体相互作用过程中产生反应性氧化物氧化,从而调节细胞质翻译。这项研究首次深入、全面地分析了植物与病原体相互作用中的氧化敏感蛋白以及核糖体蛋白氧化对作物翻译的影响,扩大了对PTM功能的理解。

