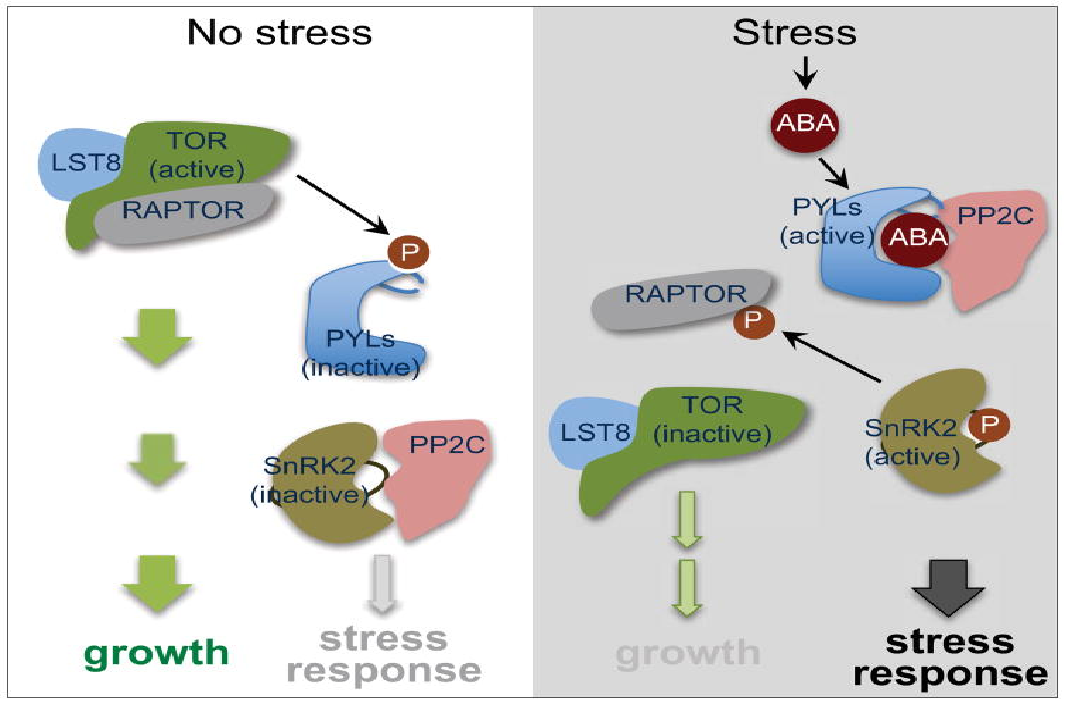
植物在环境胁迫会触发各种反应,包括由植物激素脱落酸 (ABA) 介导的生长抑制。但将压力反应与生长相结合的机制知之甚少。
中国科学院分子植物科学卓越创新中心在《Molecular Cell》期刊发布题为"Reciprocal Regulation of the TOR Kinase and ABA Receptor Balances Plant Growth and Stress Response"的文章,报道了TOR 信号在无压力条件下抑制 ABA 信号传导和应激反应,而 ABA 信号在压力时期抑制 TOR 信号和生长。植物利用这种保守的磷酸化调节反馈机制来优化生长和胁迫反应的平衡。

研究背景
在感知到环境压力时,植物会牺牲生长并激活保护性应激。植物激素脱落酸(ABA)在整合各种压力信号和控制下游压力反应方面起着关键作用。在应对干旱等环境胁迫时,ABA 迅速积累并结合PYR1/PYL/RCAR 蛋白家族(以下简称 PYL)中的受体,ABA-PYL 受体复合体会抑制 PP2C 家族分支 A 中的下游蛋白磷酸酶,PP2C 抑制会释放蔗糖非发酵-1(SNF1) 相关蛋白激酶-2 (SnRK2s),该蛋白激酶可磷酸化下游效应子以启动保护性反应,例如气孔闭合和基因表达重编程。拟南芥的PYL家族有14个成员。尽管 PYR1 和 PYL1-4 仅在 ABA 存在下与 PP2C 相互作用并抑制 PP2C,但即使没有 ABA,其余的 PYL 也可以在体外与 PP2C 相互作用并抑制 PP2C ,目前尚不清楚这些具有 ABA 独立活性的后 PYL 如何在植物中保持不活跃,以防止在没有压力的情况下 ABA 反应。
雷帕霉素靶标(TOR)是一种进化上保守的主调节因子,它整合了能量、生长、激素和压力信号,以促进所有真核生物的生长,并在调节细胞增殖、细胞大小、发育、蛋白质合成、转录和代谢方面具有重要作用 ,TOR 通过磷酸化翻译起始因子 elF3h 和 40S 核糖体蛋白 S6 激酶 (S6Ks) 来控制翻译起始,根和芽分生组织都需要 TOR 激酶 ,在根分生组织中,TOR 磷酸化 E2Fa 转录因子,以调节参与代谢、细胞周期、转录、信号转导、转运和蛋白质折叠的基因的表达,植物生长激素生长素通过 TOR 和生长素激活的 Rho 样 GTP 酶 2 (ROP 2) 之间的物理相互作用来刺激 TOR 活性,从而促进芽分生组织的激活。
技术手段
实验材料:ABA处理的拟南芥幼苗、raptor1-2 突变拟南芥、Snrk2 突变拟南芥幼苗、正常拟南芥幼苗
研究方法:磷酸化蛋白质组学、免疫共沉淀,酵母双杂、转录组学等。
研主要结果
ABA处理消除了PYL的中保守丝氨酸的磷酸化
对用和没用ABA处理的拟南芥幼苗进行磷酸化蛋白质组学,共鉴定出 47,126 个磷酸肽, 37,387 个磷酸化位点。通过数据分析发现ABA处理消除了PYL1中Ser119的磷酸化和PYL9中的Ser94(图1A),它们对应于PYL4 Ser114。作者还检测了携带天然启动子驱动的野生型 PYL1-Myc 的转基因 pyr1pyl1pyl2pyl4 (pyr1pyl124) 四突变植物免疫沉淀的 PYL1 的磷酸化,发现与前面的研究结果类似,ABA处理消除了PYL1-Myc中的Ser119磷酸化(图1C),Ser119和Ser114在拟南芥的所有14个PYLs中都是保守的。此外作者为了确定这种保守丝氨酸在PYL家族中的作用,将丝氨酸分布点突变为大小类似且不可磷酸化的中性丙氨酸 (A) 或半胱氨酸 (C),突变为更大的、不可磷酸化的中性亮氨酸 (L) 或谷氨酰胺 (Q),或突变为带负电荷的磷酸天冬氨酸 (D) 或谷氨酸 (E),ABA结合需要位于119位的小型不带电侧链(S、A、C)。Ser119 的磷酸化,使侧链扩大并带负电荷,或用较大的侧链和/或带电氨基酸(L、Q、D、E)取代,阻断 ABA 结合口袋并降低 ABA 结合亲和力。此外,PYL4 S114D 还消除了对其他分支A PP2C磷酸酶的ABA依赖性抑制,包括HAB1,PP2CA,HAI3和AHG1(图1E)。此外,在所有其他pyl的保守丝氨酸上的磷酰亚胺取代或消除都大大减少了ABA依赖的RD29B-LUC诱导(图1E)。这些结果表明,保守丝氨酸的磷酸化改变了 ABA 结合口袋并在体内负向调节 PYLs 的功能。此外作者在体外重建了核心信号通路ABA-PYL-ABI1-SnRK2.6-ABF2(脱落酸反应元件-结合因子 2)系统检查了磷酸化如何影响PYL活性,结果显示PYL1 在保守丝氨酸上的磷酸化可防止 PP2C 的 ABA 依赖性抑制(图1C、D)。为了确定保守丝氨酸的磷酸突变是否会影响 ABI1 和 PYL 蛋白之间的相互作用,作者使用了酵母双杂交进行测定(图1E)、磷酸化突变验证、互作实验(图1F)。结果显示PYLs中保守丝氨酸的磷酸化消除了与PP2Cs的ABA依赖性和ABA非依赖性物理相互作用。
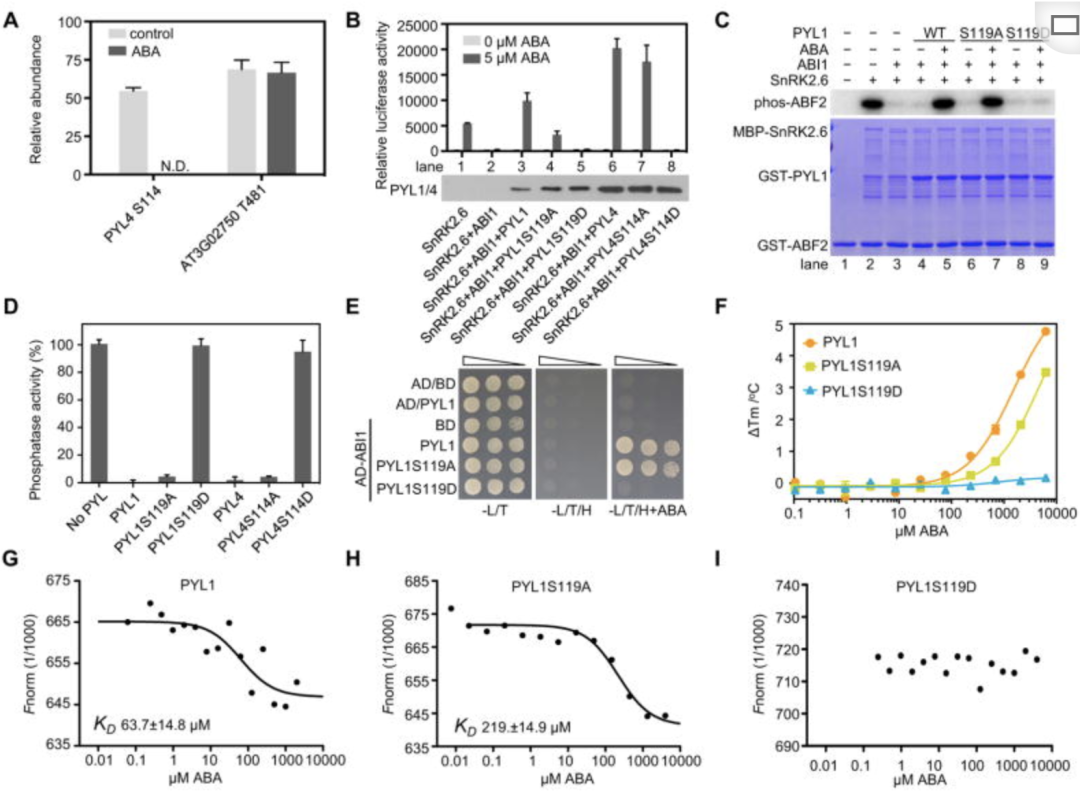
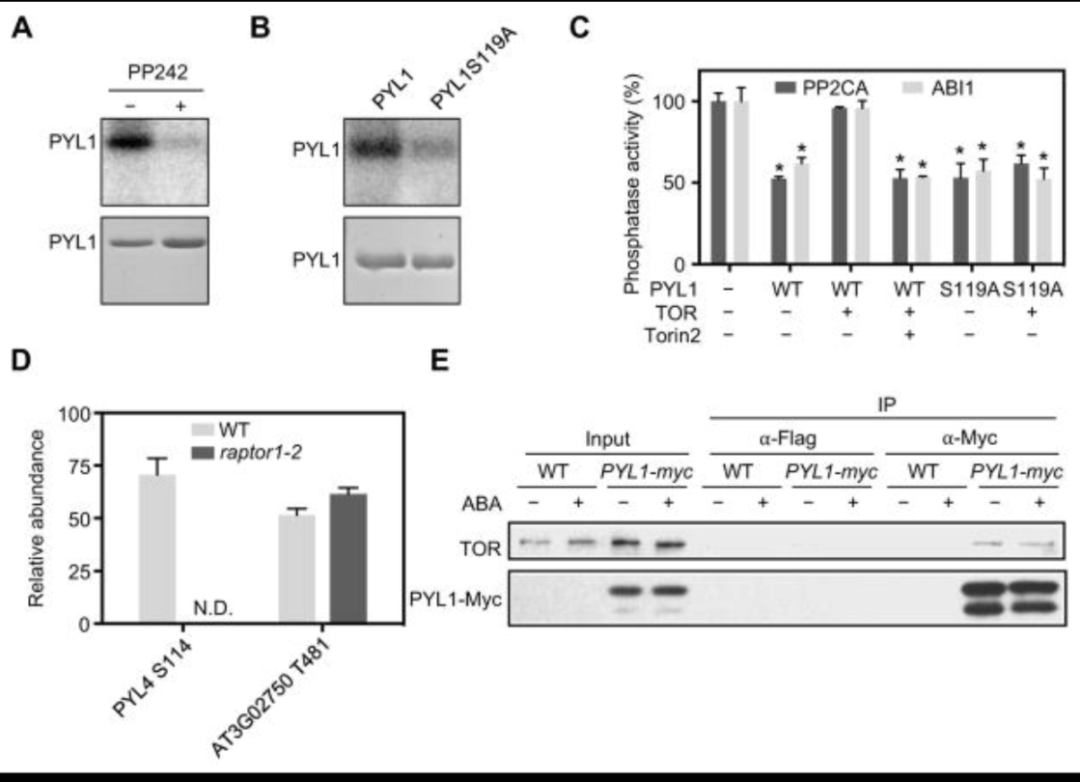
TOR 激酶在没有应激的情况下磷酸化 PYL 并负调控 ABA 信号传导
另外通过检测雌二醇诱导的TOR RNAi系和 raptor1-2 突变植物中的 ABA 反应,发现 TOR 激酶能负向调节 ABA 信号传导和应激反应;通过凝胶内测定法监测蛋白底物SnRK2活性等实验确定了TOR 或 RaptorB 功能的丧失是否会影响 ABA 信号传导;通过雌二醇处理的es-tor植物的转录组数据发现在没有应激的情况下,PYLs的TOR激酶磷酸化可阻止ABA信号传导和应激反应等。
ABA 通过 SnRK2 介导的 RaptorB 磷酸化抑制 TOR 激酶活性
作者先通过点突变、酵母双杂实验验证了 RaptorB 和 SnRK2.2,SnRK2.3,SnRK2.6和SnRK2.8之间的潜在相互作用(图6A),此外作者还发现SnRK2.6 但不是随机选择的蛋白激酶细胞周期蛋白依赖性蛋白激酶 CDKF,可以磷酸化 RaptorB 的重组片段(图 6E)。这些结果表明,RaptorB是SnRK2s的直接底物。为了进一步证实 SnRK2 活性与体内 Raptor 磷酸化的关系,作者通过磷酸化蛋白组对野生型拟南芥幼苗和Snrk2 突变植物中RaptorB蛋白的磷酸化状态进行检测分析,结果显示 Ser897是SnRK2 的一个主要的磷酸化位点,且 Ser897 的磷酸化依赖于体内 ABA 激活的 SnRK2s。
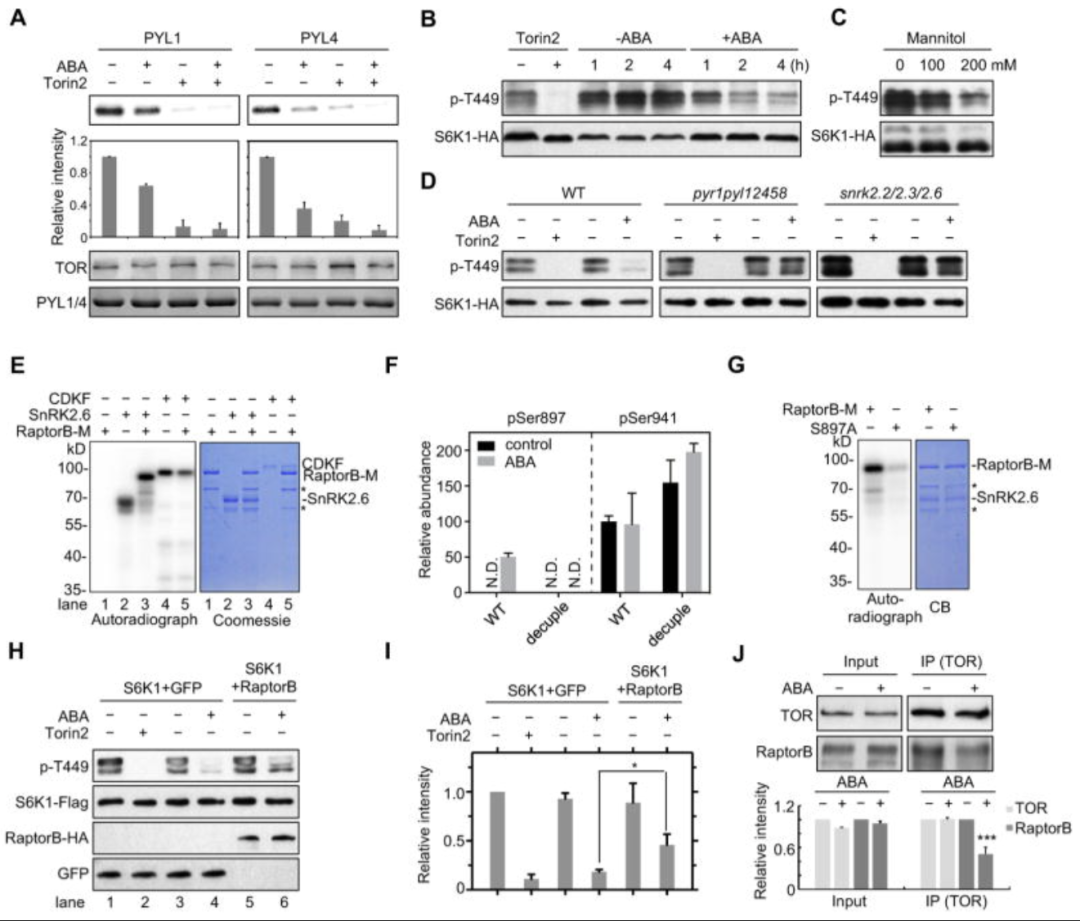
接下来,作者研究了ABA 信号转导是否可能通过影响 TOR 与 RaptorB 的关联来调节 TOR 活性,作者通过构建引入 35S 启动子驱动的 RaptorB 来提高野生型原生质体中 RaptorB 的水平,发现 RaptorB 水平的增加损害了 ABA 诱导的 TOR 抑制(图6H),此外,作者还用 ABA 处理或未经处理的幼苗中免疫沉淀了 TOR 复合物,发现ABA处理减少了RaptorB和TOR之间的相互作用(图6J),这些结果表明,ABA介导的SnRK2.6激活导致RaptorB磷酸化,从而促进RaptorB与TOR复合物的解离并抑制TOR激酶活性。
研究结论
者研究在这项研究中,作者通过对多种处理的拟南芥幼苗进行磷酸化蛋白质组组学鉴定分析,再结合磷酸化点突变、酵母双杂、免疫共沉淀等实验,发现了TOR激酶介导的PYL磷酸化代表了一种保守的机制,可在非应激条件下阻止应激信号传导,并在应激消退后使应激信号脱敏。还发现了ABA 和胁迫通过激活 SnRK2 激酶磷酸化 RaptorB(TOR 复合物的调节成分)来抑制植物生长以保存能量和确保生存的机制,PYLs的磷酸化也可以在植物的胁迫恢复过程中关闭ABA信号传导。此外,由于 PYL1 中含所含的 Ser119 的丝氨酸在 12 个不同物种鉴定都是保守的,因此 PYLs 的磷酸化调控在陆地植物中似乎是高度保守的。因而作者认为,我们认为ABA核心信号组分和TOR复合物之间的磷酸化环是植物平衡胁迫和生长反应以适应不断变化的环境的关键机制。